Shufang Yang1 ![]() ,
Bin Wu2
,
Bin Wu2
For correspondence:- Shufang Yang Email: yangshufang133494@163.com Tel:+8652382182831
Received: 9 February 2016 Accepted: 27 November 2016 Published: 21 December 2016
Citation: Yang S, Wu B. Mechanism of Huiru decoction on hyperprolactinemia in rats. Trop J Pharm Res 2016; 15(12):2641-2645 doi: 10.4314/tjpr.v15i12.15
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the anti-hyperPRL mechanism of traditional Chinese medicine Huiru Decoction (HD) in rats.
Methods: The effect of HD on serum prolactin (PRL), estradiol (E2), progesterone (PGN), follicle stimulating hormone (FSH) and luteinizing hormone (LH) levels were investigated in hyperprolactinemia (hyperPRL) rats. Furthermore, the effect of HD on hypothalamus dopamine D2 receptor, protein kinase A (PKA) and cyclic adenosine monophosphate (cAMP) levels of hyperPRL rats were investigated respectively.
Results: Compared with the model group, a high dose of HD (19.2 g/kg body weight) and medium dose of HD (9.6 g/kg body weight) reduced PRL level of hyperPRL rats effectively (p < 0.01). HD increased dopamine D2 receptor (p < 0.01), reduced cAMP (p < 0.01) and PKA (p < 0.01) levels of hypothalamus in hyperPRL rats significantly.
Conclusions: HD showed anti-hyperPRL activity via dopamine D2 receptor.
Introduction
Hyperprolactinemia, which is one of the most common endocrine disorders of the hypothalamus-pituitary axis (PRL > 25ng/mL) in young women, is associated with galactorrhea and ovulatory dysfunction that results in menstrual irregularities [1]. HyperPRL can occur at any age, and the prevalence varies from 0.4 % in the normal adult population to as high as 9-17 % in women with menstrual problems such as amenorrhea or polycystic ovarian syndrome [2,3]. Common causes which induce hyperPRL are hypothalamus-pituitary lesions, pituitary tumor, severe liver or kidney disease, neuritis or irritations of the spinal cord, depression or other physiological factors such as pregnancy and lactation [4,5]. Galactorrhea is a common kind of female disease induced by hyperPRL. Synthetic drugs are used in treating them, but they always bring many side effects such as menstrual disorder and the relapse rate is very high. Bromocriptine and cabergoline are effective in curing hyperPRL, but 12 % patients cannot endure bromocriptine [6].
Empirical evidence suggests that many herbal medicines possess the therapeutic potential to alleviate hyperPRL symptoms [8]. In China, Huiru Decoction (HD) is composed of Chai Hu (Bupleurum chinense DC.) 60 g, Xia Ku Cao (Prunella vulgaris L.) 15 g, Xiangfu (Cyperus rotundus L.) 12 g, Danpi (Tree Peony Bark) 10g, Gancao (Glycyrrhiza uralensis Fisch) 10g and Dan Shen (Salvia miltiorrhiza Bge.) 10 g. It is often used to treat galactorrhea effectively without toxic side effect. In previous study, we found that HD could decrease prolactin level in hyperPRL mice effectively [9]. However, the anti-hyperPRL mechanism of it was still unknown.
In order to clarify the mechanisms underlying the anti-hyperPRL effects of HD, we planned a direct experimental test. Most conventional anti-hyperPRL agents reduce PRL secretion through D2 receptor agonism in the hypothalamic neuroendocrine dopaminergic system, and cAMP/PKA signal transduction pathway is one of dopamine receptor classic pathways [10]. And other sex-steroids other than PRL and DAT are also involved in the pathophysiology of hyperPRL [11]. Therefore, we hypothesized that the therapeutic efficacy of HD in alleviating hyperPRL could be attributed to modulation of dopamine D2 receptor. To test this hypothesis, we examined the effects of HD on dopamine D2 receptor, PKA and cAMP levels in a rat model of hyperPRL.
Methods
Preparation of HD
The dried herbs of HD were decocted with 1000 ml water twice, 45 min for each. Then the liquor was filtrated by a funnel and concentrated into 200 ml of HD. The HD was used for animal experiments.
Experiment in animal model of hyperPRL
Female Wistar rats weighing 200-220 g were obtained from Jiangsu Center for Disease Control and Prevention, Nanjing, Jiangsu. The animals had free access to food and water, and were allowed to acclimatize for at least one week before use. All experiments were approved by the Animal Care and Use Committee and were carried in compliance with the Animal Welfare Act and the NIH guidelines (NIH publication No.80-23, revised 1996).
Rats were given intraperitoneal (i.p.) of metoclopramide (MCP, 150 mg/kg daily) for 10 days to prepare hyperPRL model [12]. 60 rats were divided into six groups of ten individuals: control, model group, model plus positive drug bromocriptine group, plus high dose of HD (19.2 g/kg) group, plus middle dose of HD (9.6 g/kg) group and plus low dose of HD (4.8 g/kg) group. Each dose was dissolved in 2 mL of water, and given into rats by administered gavage. The dosage was calculated from the daily human AFH clinical dosage based on body surface area. Control and model rats received 2 ml of water. All group mice were intragastric administrated for 30 days.
Sex hormones determination
After the treatment of administrating for 30 days in hyperPRL rats, serum was obtained by eyeball removal. Serum PRL, E2, PGN, FSH and LH levels of the rats were measured by ELISA kits (Shenzhen Xin-Bo-Sheng Biological Technology Co., Ltd., China).
Western blot analysis of hypothalamus dopamine D2 receptor
Rat hypothalamus tissues were homogenized in 10 wt/vol RIPA buffer with protease inhibitor and centrifuged at 3000×g for 15 min at 4 oC. The supernatant samples were centrifuged at 12,000 × g for 20 min at 4 oC. The final peptides were dissolved in a RIPA buffer and loading buffer. After resolution of hypothalamus protein (equal loading for each sample) by 12 % sodium dodecyl sulfate polyacryla-mide gel electrophoresis, the protein samples were transferred onto polyvinylidene difluoride membranes (Merck), respectively. Nonspecific protein-binding sites were blocked with phosphate-buffered saline containing 0.1 % Tween-20 and 5 % fat-free milk for 1 h at room temperature, and then incubated in appropriate primary antibodies (DRD2, 1: 1000; ß-actin, 1: 5000). Immunoreactive bands were visualized by incubation with lumiGLO reagent (Cell Signaling) and exposed to X-ray film (Kodak, Stamford, CT, USA). Quantification of Western blot analysis was performed with ImageJ (National Institutes of Health, Bethesda, MD, USA). ß-actin levels were used as a protein loading control. The relative levels of target proteins were determined after normalization with ß-actin, respectively.
Measurement of hypothalamus PKA and cAMP levels
Rat brains were stripped and grinded into homogenate, then centrifugated for 15 min at 3000 r·min-1. The supernatant was used to measure PKA and cAMP levels by ELISA kits (Shenzhen Xin-Bo-Sheng Biological Technology Co., Ltd., China).
Statistical analysis
Values are expressed as mean ± SEM. Significant differences between the groups were analyzed using one-way analysis of variance (ANOVA) followed by two-paired Student’s t-test. P < 0.05 was considered statistically significant.
Results
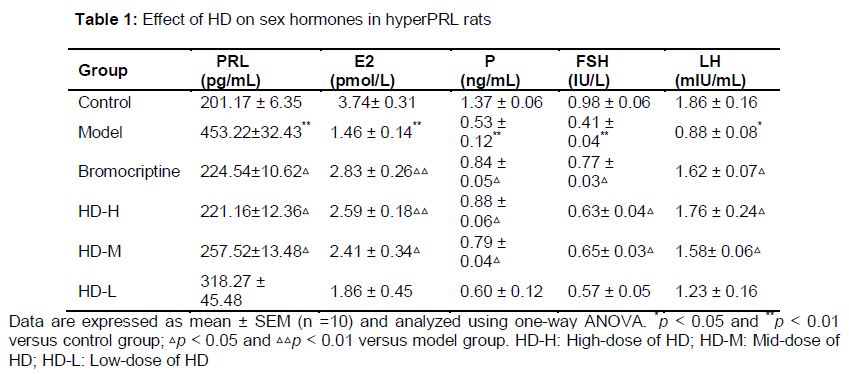
Effect of HD on sex hormones in rat model of hyperPRL
Compared with control group, serum PRL level of hyperPRL model rats increased significantly (p < 0.01). Serum E2, PGN and FSH and LH (p < 0.05) levels of hyperPRL model rats decreased significantly (p < 0.01). Compared with model group, the increased PRL level was significantly attenuated by treatment with 0.6 mg/kg bromocriptine and 9.6 or 19.2 g/kg HD after 30 days (p < 0.01). And rat serum E2 level decreased (p < 0.01), and serum PGN, FSH and LH levels increased (p < 0.05) effectively in bromocriptine-treated group. Compared with model group, rat serum E2 level decreased (p < 0.01), and serum P, FSH and LH levels increased (p < 0.05) significantly treated by high-dose of HD ().
Effects of HD on hypothalamus dopamine D2 receptor in rat model of hyperPRL
Western blot analysis revealed that hypothalamus dopamine D2 receptor protein expression in model rats was much less than that in control rats (p < 0.01). Compared with model rats, such decreased dopamine D2 receptor protein were significantly improved by treatment with 0.6 mg/kg bromocriptine (p < 0.01) or 4.8 g/kg (p < 0.05) or 9.6 g/kg (p < 0.01) or 19.2 g/kg (p < 0.01) HD for 30 days.
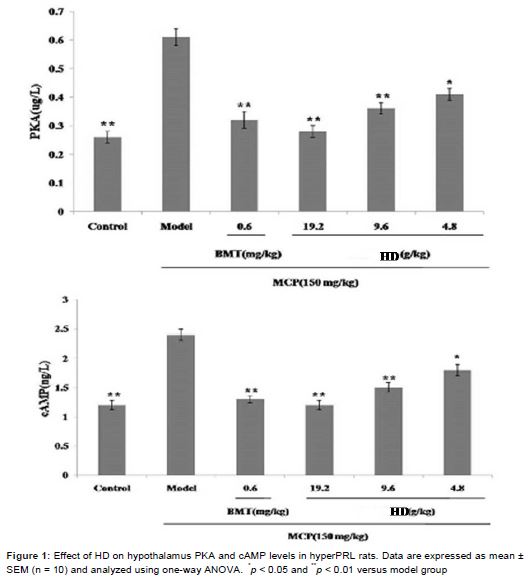
Effect of HD on hypothalamus PKA and cAMP levels in rat model of hyperPRL
Our study demonstrated that hypothalamus PKA (p < 0.01) and cAMP (p < 0.01) level in rat model of hyperPRL increased significantly compare to control group. Such increased PKA and cAMP were significantly attenuated by treatment with 0.6 mg/kg bromocriptine or 4.8 g/kg (p <0.05) or 9.6 g/kg (p < 0.01) or 19.2 g/kg (p < 0.01) HD for 30 days ().
Discussion
In previous study, we found that HD could decrease prolactin level in hyperPRL rats effectively. In this study, HD increased dopamine D2 receptor, reduced the hypothalamus cAMP and PKA levels in hyperPRL rats significantly.
Dopamine receptors belong to the family of seven transmembrane domain G-protein coupled receptors (GPCR) [13-15]. Dopamine receptors D1 and D2 are classified into two subfamilies based on their differential effect on adenylyl cyclase.
Classically, the functions of dopamine receptors have been associated with the regulation of adenylate cyclase-protein kinas A (cAMP–PKA) through G-protein-mediated signaling. Two classes of GPCR mediate dopamine functions, D1-like receptor subtypes (D1 and D5) couple mostly to Gas and stimulate the production of the second messenger cAMP and the activity of PKA. However, D2-like subfamily (D2, D3, and D4) couple to Gai/o and regulate the production of cAMP negatively, thus resulting in a diminution of PKA activity [16-18].
The physiological and pathological roles of DR2 have been recognized in some organs such as brain and kidney [19,20]. In the adenohypophysis, the predominant dopamine receptor is the D2 receptor [21]. Transfection of the dopamine D2 receptor into a pituitary cell line results in a decrease in intracellular cAMP and inhibits prolactin secretion when dopamine is added to the cell culture [22,23]. Most conventional anti-hyperPRL agents reduce PRL secretion through D2 receptor agonism in the hypothalamic neuroendocrine dopaminergic system, and cAMP/PKA signal transduction pathway is one of dopamine receptor classic pathways. In our study, HD inhibited PRL level via dopamine D2 receptor, and it reduced the cAMP and PKA levels of hypothalamus in hyperPRL rats significantly. These suggested that HD showed anti-hyperPRL activity via dopamine D2 receptor.
Conclusion
Our studies demonstrated that HD show anti-hyperPRL activity via dopamine D2 receptor.
References
Archives


News Updates